Nutrition and diet
A review of folate status in diet and DNA methylation with an explanation by nutrient-gene interaction
Abstract
Recently an interaction was found between folate and DNA methylation, explaining an impaired epigenetic control of gene expression due to a deficiency of folate in diet. Studies based on animal models have shown that this deficiency leads to mutation in Methylenetetrahydrofolate reductase that affects the one carbon metabolism. MTHFR polymorphism at 677 base pair leads to less production of S — adinosyl methionine (SAM), the universal methyl donour. Less SAM results in inadequate DNA methylation and impaired nucleotide synthesis. At the same time, being a co-enzyme and substrate in the conversion of homocysteine to methionine, the mutation of MTHFR interrupts the entire process, leading to increased levels of homocysteine in blood which results in CVDs. On the other hand, the thymine-thymine genotype of MTHFR (MTHFRTT) is also found to act in carcinogenesis. These findings which relate to the relationship between plasma folate levels and MTHFR polymorphism not only explain development of certain diseases but also open a new area of ‘Individualised nutrition’ where prevention can be taken by modifying existing diet.
Introduction
Nutrigenomics, a branch of nutrition science, seeks to provide the mechanism of how a specific nutrient interacts with genes to control their expression. Nutrition research has recently highlighted the role of several nutrients in regulating the genome machinery. The micro-nutrients especially play an integral role in genetic modulation, synthesis, expression and repairing the system. Folate is such a nutrient that modulates DNA methylation. By gaining knowledge on specific mechanism of DNA methylation by folate we can now make an approach towards ‘personalised nutrition’ for better health and lifestyle (1).
What is folate?
Folates are important co-factors in the transfer and utilisation of one-carbon moieties and play a key role in the synthesis of nucleic acids and methionine generation. Folate deficiency causes a defect in DNA synthesis, causing megaloblastic macrocytic anaemia. Folate deficiency also causes intra-uterine-growth-retardation, a neural tube defect in the foetus, developmental abnormalities etc (2, 3).
What is DNA methylation?
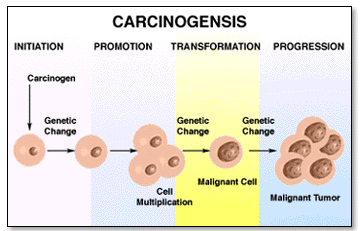
DNA methylation, a characteristic feature of many eukaryotic genomes, consists of the addition of a methyl group at the carbon 5' position of the cytosine within the cytosine-guanine (CpG) dinucleotide in a complex reaction that probably involves the flipping of the cytosine base out of the intact double helix. Typically, DNA methylation occurs in CpG-di-nucleotide rich regions (CpG Island) (4). DNA methylation is the fundamental mechanism for the epigenetic control of gene expression and the maintenance of genomic integrity (5, 6). Therefore, an evaluation of DNA methylation status is important for the study of cell growth regulation, tissue specific differentiation (2, 4, 7) and carcinogenesis (6).
Several nutrients and their effects on genetic integrity and methylation
Reduced dietary intake or low plasma level of several nutrients have been associated with faulty metabolic pathways, developing cancer and abnormal gene expression (1, 8).
Based on animal studies, it is established that many nutrients, especially vitamins and minerals, are essential for maintaining genomic stability. For example:
• Vitamin C and Vitamin E deficiencies are known to cause DNA oxidation and chromosome damage (9, 10).
• Being an antioxidant, Vitamin D has been found to stabilise chromosomal structure and prevents DNA double strand breaks (11).
• A carotene-rich diet has been shown as to reduce DNA damage (12).
• Vitamin B12 deficiency is associated with micro-nutrient formation (1, 13).
The studies provide an idea of how the mechanism of nutrient and gene interaction is particularly important in maintaining genomic integrity and expression.
Effect of folate deficiency on DNA methylation — an explanation of gene-nutrient interaction
Folate deficiency and related outcome can be explained under the three following headings:
• Less production of S-adenosylmethionine (SAM)
Folate, in its co-enzyme form is involved in DNA methylation by one-carbon metabolism. The sole function of all co-enzymes is to transfer one-carbon units. Within this scope, SAM is synthesized by these co-enzymes which are known as the universal donor of methyl group within the body. It is also important in the synthesis of denovo-deoxynucleotide triphosphate synthesis. The reduced availability of methyltetrahydrofolate (MTHF) decreases the biosynthesis of SAM, thus limiting the availability of methyl group for DNA methylation. Thus, dietary folate depletion leads to inadequate DNA meth-ylation both in humans (14, 15) and animals (16).
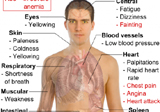
• Less production of methionine from homocysteine — increased risk of CVD (cardio-vascular diseases)
Methionine is regenerated from homocysteine by methionine synthase in a reaction where 5–methyl tetrahydro folate (5-MTHF) acts both as a co-factor and substrate. The reduced level of MTHF leads to impaired conversion of methione from homocysteine. Elevated level of plasma homocysteine increases the risk of CVDs. Epidemiological as well as experimental studies have shown that folate deficiency is a major cause of hyperhomocysteinemia which is fully recognised as an independent risk factor for atherosclerosis (17).
Two recent studies have demonstrated that folic acid supplementation in patients with asymptomatic hyper-homocysteinemia can improve endothelial function, measured as enhanced flow-mediated vasodialation (18).
Mechanism of action
Methyl tetrahydrofolate reductase (MTHFR) is considered as the key enzyme in one-carbon metabolism because it catalyses the irreversible conversion of (5, 10) — methyl tetrahydrofolate to methyl tetrahydrofolate (MTHF) (17). In 1988, Kang and colleagues identified a variant of the MTHFR that causes enzyme thermolability and reduced activity (18). This mutant enzyme was associated with elevated plasma homocysteine levels, which is to be expected because the conversion of homocysteine to methionine is impaired. The thermo-labile variant of the MTHFR is due to a common missense mutation, a cytosine-to-thymine transition at base pair 677 (C677T) (19) that results in an alanine-to-valine substitution in the MTHFR amino acid sequence.
• MTHFR and carcinogenesis
The mutant thymine/thymine (TT) genotype of MTHFR polymorph is associated with a lower risk of causing colorectal cancer; however this protective factor is observed only in persons with adequate folate status. But, persons with lower level of folate in blood show higher risk of developing colorectal cancer even with the presence of TT genotype mutant gene (20). Distribution of different co-enzyme forms of folate is altered in MTHFRTT homozygotes (21). The RBCs of TT homozygous mutants show variable amounts of formylated tetrahydro-folate polyglutamates instead of methylated tetrahydrofolate; whereas, cells from the cytosine-cytosine (CC) wild type persons contain exclusively methylated tetrahydro-folate derivatives (21).
Folate and MTHFR gene interaction
Gemonic DNA methyltion is dependent upon the gene-nutrient interaction between the MTHFRC677T variant and folate level in plasma. The enzyme MTHFR is responsible for the irreversible conversion of 5, 10 — methylhydrofolate to 5-methyltetrahydro-folate (8). The mutant MTHFRC677T encodes for thermolabile enzyme with lower activity. Persons carrying in homozygosity the mutant allele for the thermolabile MTHFR (TT) have a decreased formation of MTHF, resulting in less production of S-adenosyl methionone, the constant supplier of — CH3 for different methylation reactions, including DNA methylation (9). On contrary, subjects with MTHFR (CC) genotype are not affected by folate deficiency, as the synthesis of MTHF for methylation reactions and conversion of homocysteine to methionine are preserved. The MTHFR doesn’t alter the availability of CH3-THF when folate status is adequate.
Inclusion of folate in diet — an approach to ‘personalised nutrition’
The review discusses recent data in the field of gene-nutrient interactions and DNA methylation, a fundamental epigenetic feature of DNA that affects gene expression and genome integrity. Several nutrients affect DNA metabolism, repair and synthesis and folate is an essential vitamin among them. The review discusses effect of folate deficiency on MTHFR mutation, a key enzyme of the one-carbon metabolic pathway and methylation. It also describes the mechanism of methionine production from homocysteine by one-carbon metabolic reactions and related CVDs. ‘Personalised nutrition’ will allow one to choose better food, enriched with folate that will lead to proper brain development, DNA synthesis and ultimately sound mental health.
What high folate foods can do for us
• Support RBC production and help prevent anaemia
• Help to prevent homocysteine build-up in the body
• Support skin cell production
• Accelerate methionine production
• Prevent dementia and Alzheimer’s disease
Food sources
• Lentils
• Dark-green leafy vegetables
• Beans
• Spinach
• Turnip greens
• Broccoli
• Cauliflower, etc.
References
1. French M, Ferguson LR. Vitamins/minerals and genomic stability in humans. Mutat. Res. 2001; 4759 (13): 1-6.
2. Di Bello PM, Dayal S, Kaveti D, Zhang D, Kinter M, Lentz SR, et al., The nutrigenetics of hyperhomocysteinemia: quantitative proteomics reveals differences in the methionine cycle enzymes of gene-induced versus diet-induced hyperhomocysteinemia, Mol. Cell. Proteomics 2010; 9 (4): 471-85.
3. Robertson KD, Wolffe AP. DNA methylation in health and disease. Nat. Rev. Genet 2000; 1: 11-9.
4. Wolffe AP. Matzke MA. Epigenetics: regulation through repression. Science 1999; 286 (7): 481-86.
5. Lister R, Pelizzola M, Dowen RH, Hawkins RD, Hon G, Tonti-Filippini J, et al. Human DNA methylomes at base resolution show widespread epigenominc differences. Nature 2009; 462 (6): 315-22.
6. Feinberg AP. Methylation meets genomics. Nat. Genet. 2001; 27(3): 9-10.
7. Da Costa K, Badea M, Fischer L, Zeisel S. Elevated serum creatine phosphokinase in choline-deficient humans: mechanistic studies in C2C12 mouse myoblasts. Am J Clin Nutr 2004; 80(3): 163-70.
8. Ames R, Halliwell B. Vitamin C and genomic stability. Mutat. Res. 2001; 475(2): 29-35.
9. Claycombe KJ, Meydani SN. Vitamin E and genome stability. Mutat. Res. 2001; 475(2): 37-44.
10. Zeisel SH. Genetic polymorphisms in methyl-group metabolism and epigenetics: lessons from humans and mouse models. Brain Res. 2008; 1237(2): 5-11.
11. Collins AR. Carotenoids and genomic stability. Mutat. Res. 2001; 475: 21-8.
12. Teschendorff AE, Menon U, Gentry-Maharaj A, Ramus SJ, Weisenberger DJ, Shen H, et al. Age-dependent DNA methylation of genes that are suppressed in stem cells is a hallmark of cancer. Genome Res. 2010; 20 (3): 440-6.
13. Zschäbitz S, Cheng TY, Neuhouser ML, Zheng Y, Ray RM, Miller JW, et al. B vitamin intakes and incidence of colorectal cancer: results from the Women’s Health Initiative Observational Study cohort. Am J Clin Nutr 2013; 97(6): 332-43.
14. Fazzari MJ, Greally JM. Introduction to epigenomics and epigenome-wide analysis. Methods Mol Biol. 2010; 620(4): 243-65.
15. Clin chem lab Med. 1988 Jun; 36(7): 419-29. [Online] Available from: www.ncbi.nlm.nih. gov/Pubmed/9746265 [Accessed 10th June 2012].
16. Woo KS, Chook P, Lolin YI, Sanderson JE, Metreweli C, Celermajer DS, et al. Folic Acid improves arterial endothelium functions in adults with hyperhomocysteinemia, J Am Coll Cardiol. 1999; 34(3): 2002-6.
17. Roger VL, Go AS, Lloyd-Jones DM, Adams RJ, Berry JD, Brown TM, et al. Heart disease and stroke statistics — 2011 update: a report from the American Heart Association. Circulation 2011; 123 (11): 18-209.
18. Bailey LB. Folate, methyl-related nutrients, alcohol, and the MTHFR 677CàT polymor-phism affect cancer risk: intake recom-mendations. J Nutr 2003; 133: 3748S-53S.
19. Blom HJ, Smulders Y. Overview of homo-cysteine and folate metabolism. With special references to cardiovascular disease and neural tube defects, J. Inherit. Metab. Dis. 2011; 34(4): 75-81.
20. Chang SC, Lin PC, Lin JK. Role of MTHFR polymorphisms and folate levels in different phenotypes of sporadic colorectal cancers. Int J Colorectal Dis 2007; 22(8): 483-9.
21. Gallegos-Arreola MP, Garcia-Ortiz JE, Figuera LE. Association of the 677C ->T polymorphism in the MTHFR gene with colorectal cancer in Mexican patients. Cancer Genomics Proteomics 2009; 6(5): 183–8.
Ms. Subhasree Ray is Nutritionist of Nutrition Rehabilitation Centre (NRC), Birbhum, Department of Health & Family Welfare, Govt. Of West Bengal and Research Scholar at SNDT University, Mumbai, India.
Share with us (Comments,contributions,opinions)
When reproducing this feature, please credit NAMAH,and give the byline. Please send us cuttings.